随着人类活动和全球气候变化的加剧,全球性物种灭绝的速度正在加快。尽管在1992年巴西里约热内卢召开的联合国环境与发展大会上,153个国家签署了《保护生物多样性公约》,然而,收集的数据显示,截止到2010年,全球尺度生物多样性丧失仍在继续,且速度有增加的趋势[1]。在越来越趋于简化的生态系统中,生物多样性如何影响生态系统功能成为生态学家最为关注的重大科学问题之一。系统属性随着时间波动是其自然特性之一,即使在环境因子不变的情况下,这种波动性依然存在[2]。波动性加剧(稳定性下降)说明系统受到的压力增大[3],将会增加物种灭绝的风险[4-5],使生态系统过程更加难以理解和预测[6],甚至使生态系统提供产品和调节气候等功能彻底丧失[7],因此在诸多生态系统功能中生态系统稳定性备受关注。
生态系统稳定性的影响因子众多,非生物因子包括气候(温度,辐射,降水等),资源的可获得性(N、P的浓度等)以及干扰(施肥,火烧等);生物因子包括生物多样性,物种相互作用的强度,以及食物网的拓扑结构[2, 8-9],其中生态学家对生物多样性的关注更多[10]。这是因为全球性生物多样性正在锐减,除此以外,还因为生物多样性具有以下特点: 1) 生物多样性便于操控和测量[2],例如Ceedar Creek草地试验中用7种不同的氮添加量来控制自然生态系统的物种丰富度[11],欧洲草地实验(BIODEPTH)以人工播种的方式在8块草地上控制物种丰富度[12];2) 生物多样性对生态系统功能的作用强度与其它影响因子相当,最近通过对11个实验的整理分析和192篇相关文献的结果进行整合分析(meta-analysis),生态学家发现生物多样性是驱动生态系统过程的关键因子[9, 13];3) 生物多样性对任何系统功能的影响都是非线性的饱和形式[14],这意味着,初期的生物多样性丧失对生态系统功能的影响略小,在物种数目低于某一阈值后,任何物种的灭绝都会对生态系统功能产生严重的影响,对于这一阈值的定量化研究具有重要的生态学意义。此外一些研究表明如果考虑更长的时间尺度、更大的空间尺度、多个系统功能以及复杂的环境事件,曲线将很难达到饱和[15-17],说明在更大的时空尺度下,系统中的任何物种可能在某一特定环境事件中,对一个或多个系统过程起关键作用,这些研究对物种冗余的假说产生了质疑[18]。总之,由于生物多样性的重要意义和特殊性质,生物多样性相关研究一直吸引着无数生态学家的研究热情,也使得近60年来生物多样性与稳定性关系备受关注且争议不断。
本文首先对生物多样性稳定性研究的历史进行简单的回顾,然后根据时间稳定性的计算方法将生物多样性稳定性机制划分为提高表现效应,方差之和效应和协方差之和效应。在此基础上对现在生物多样性与稳定性研究中最常见的机制进行详细介绍,然后对生物多样性与稳定性研究中存在的主要争议进行总结并对未来生物多样性研究进行展望。
1 生物多样性与稳定性研究的回顾许多生态学家追溯过生物多样性与稳定关系的研究起源,Tilman[8]认为Clemen[19] 1916年和Smuts[20] 1926年的著作对随后的生物多样性与稳定性研究有所启发,Jiang 与 Pu[10]通过查阅资料认为"生物多样性可能影响稳定性"的提法最早见于Elton[21] 1927年的著作。随后,到20世纪50年代,Odum,MarArthur以及Elton的工作真正拉开了生物多样性稳定性研究的序幕。Odum[22] 通过重复的观察认为种群密度在简单陆地群落中比在复杂群落中更易发生剧烈波动。MarArthur[23] 解释了捕食者和被捕食者数量丰富的群落在受到干扰后,种群的多度在变化后可以迅速恢复。Elton[24]表明多样性高的群落抵抗外来种入侵的能力更强。
20世纪70年代,Gardner 与Ashby[25] 和May[26]等开始用数学模型的方法严格的检验生物多样性与稳定性的关系,他们发现与前人的研究结果相反,生物多样性使系统的稳定性下降。Pimm 与 Lawton[27]和Yodzis[28]用类似的方法得出了相同的结论。Goodman[29]在1975年对当时已发表的超过200篇相关文献进行综述,发现当时的证据并不能证明生物多样性稳定性假说。在随后接近20年的时间里,生态学家并没有发现生物多样与稳定性间存在明显关系。
直到20世纪90年代中期,生态学家开展了一系列生物多样性试验,其中代表性的实验包括生态箱实验、微宇宙实验、Cedar Creek草地多样性实验和欧洲草地实验[30],这些实验的开展让人们看到一个生物多样性研究的全新时代的到来。其中Tilman 与 Dowing[11] 1994年报道的Cedar Creek草地多样性试验结果以及Naeem 与Li[31] 1997年报道的微宇宙实验的结果重新点燃了生态学家对生物多样性和稳定性的研究热情。
最近十几年有关生物多样性与稳定性关系的研究持续增长[32]。更多的生物多样性实验在淡水、海洋、陆地等生态系统展开,据统计到2009年有数百篇的相关文献报道了超过600个生物多样性实验的研究结果[14]。这些实验对已有的理论进行检测的同时为新理论的提出奠定了实验基础。此外,许多学者用数学模型研究生物多样性与稳定性的关系[33-40],他们的工作为新理论的提出做出了突出的贡献。部分生态学家在前人的工作基础上对已有成果进行理论综述,如Tilman[8],McCann[41],Cottingham等[5],Ives 与 Carpenter[2],Jiang 与 Pu[10],Loreau 与de Mazancourt[7]等人的研究,这些工作使生物多样性稳定性的研究更具系统性,同时对人们了解生物多样性稳定性关系研究进展和理解生物多样性与稳定性关系的内在机制有重要帮助。
2 稳定性计算稳定性在生态学中有广泛的定义,Pimm[4, 42]给出了45个稳定性变量,其中常见的变量包括:抵抗力,恢复力,持久性和变异性。抵抗力和恢复力是从系统抵抗变化的角度考虑,持久性和变异性则从系统内部动态的角度考虑[2, 41]。由于不同的定义会导致不同的生物多样性稳定性关系,因此在研究中声明所采用的稳定性定义非常必要[43]。在经验和理论研究中,生态学家多采用变异性来衡量群落稳定性[41],因此本文以下主要关注变异性和与变异性密切相关的时间稳定性。传统上,人们通常用变异系数来测量变异性,因此时间变异性可以用一段时间内多度平均值μ和标准差σ来表示[33]即:CV=100σ/μ。在此基础上Tilman[8]将时间稳定性定义为S=μ/σ。从群落组成的角度,群落变异性可以表述为[5, 44]:
 (1)
(1)
式中,Xc是群落的属性(例如生物量或多度),是群落中所有物种属性值之和,Xi是物种i的属性,Var(xi)群落中物种i属性的方差即变异值,Cov(xi,xk)是物种i和物种k的属性的协方差。该公式的生态学意义为群落多度的方差为所有组成物种的方差之和与成对物种间协方差之和的累加,由于标准差为方差的平方根,因此时间稳定性还可以表述为[43]:
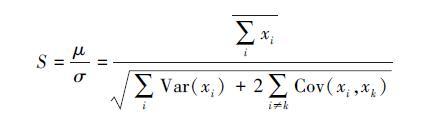 (2)
3 生物多样性对稳定性的作用机制
(2)
3 生物多样性对稳定性的作用机制 随着生物多样性与稳定性研究数量的迅速增加,一系列生物多样性与稳定性关系的内在机制被提出。这些机制往往从不同角度考虑,通过不同的方法提出,且机制间并不相互独立,因此使初步接触该领域的人们感到困惑。通过公式(2)可以将时间稳定性划分为平均值、方差之和以及协方差之和3个统计组成部分。生物多样性可以通过提高表现效应,方差之和效应和协方差之和效应改变群落的时间稳定性[10, 45-46]。本文按图 1对如今生物多样性与稳定性研究中最常见的机制进行分类并对各机制展开详细说明。
3.1 提高表现效应-超产超产是指多个物种混播的群落比其任何组成物种单播的生产力高[47]。如果生物多样性的提高只引起平均值的增加,对其他变量不产生影响,那么更高的生产力将会导致更高的时间稳定性[8]。超产因此被认为是生物多样性与稳定性关系内在机制之一。许多生物多样性实验发现超产现象的存在,更多的实验发现随着生物多样性的提高群落的生产力在增加[14-15, 48-50]。超产是一个有趣的现象,其发生机制引起了生态学家的浓厚兴趣。其中选择效应和互补效应与超产的产生密切相关[46]。选择效应认为具有特殊性质的物种在群落占优势会影响生态系统过程。互补效应认为不同的物种具有特定的生态位,生物多样性的增加有助于物种之间的功能互补,使群落捕获更多的资源进而引起超产[51]。Cardinale等[50]对44个生物多样性实验进行整合分析表明,选择效应解释了三分之一的生物多样性效应,而互补效应解释了三分之二。
许多研究表明超产是解释生物多样性与稳定性关系的重要机制。Loreau 与 de Mazancourt[7]通过Lotka-Volterra竞争模型将环境的波动性和种群动态的随机性相结合,证明超产在样地水平或种群较小时将会起到比较重要的作用,而现在的生物多样性实验大多为这种情况。Jiang 与 Pu[10]对29个研究进行整合分析,结果表明超产比方差之和效应和协方差之和效应更能解释生物多样性与稳定性间的正相互作用。Hector等[46]对欧洲草地实验(BIODEPTH)的分析结果显示随着生物多样性水平的增加,平均值的增加程度比标准差的增加程度大,因此超产是该实验中重要的稳定性机制。de Mazancourt等[40]的理论模型重点关注植物内在性质,通过该模型对包括Cedar Creek实验在内的4个长期草地生物多样性实验进行检验,发现4个实验中,超产在维持群落稳定性方面均发挥了一定的作用。
3.2 方差之和效应自然群落由不同的物种组成,这些物种彼此间相互作用,同时它们的多度也在随时间而发生随机波动。如果假设物种彼此相互独立,则物种协方差之和为零,因此引起方差之和变化的一些机制必然在生物多样性与稳定性关系中发挥作用。
3.2.1 投资组合效应投资组合效应是一个经济学概念,是指多样化的投资组合更加稳定。纯粹的从统计学角度出发,假设物种之间彼此相互独立,则物种的协方差之和为零。那么公式(1)可以表述为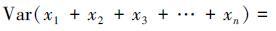
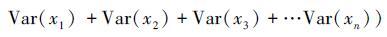 。用标准差的形式可表述为
。用标准差的形式可表述为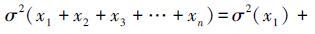
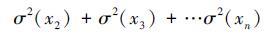 。假设群落的总多度μ不随物种数发生改变,且群落中物种的多度和方差相同[33],那么
。假设群落的总多度μ不随物种数发生改变,且群落中物种的多度和方差相同[33],那么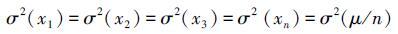 。单物种群落的时间稳定性为:Ss=μ/σ(μ),多物种群落的时间稳定性为
。单物种群落的时间稳定性为:Ss=μ/σ(μ),多物种群落的时间稳定性为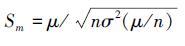 ,tp官方正版下载因为
,tp官方正版下载因为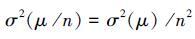 ,
token钱包安卓版下载所以
,
token钱包安卓版下载所以
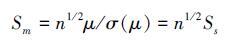 (3)
(3)
可见群落的时间稳定性随物种数的增加而增加(图 2)。由于σ2(μ/n)= σ2(μ)/n2的存在,Doak等[33]认为投资组合效应的存在是不可避免的。这种用数学运算来解释生物多样性与稳定性之间的关系与经济学中货币存量的增减与资产结构间的关系相似,由生态学家引入称为投资组合效应[52]。
3.2.2 均匀度和优势种的稳定性均匀度往往被认为通过丰富度直接或间接影响群落的稳定性[53]。在投资组合效应的基础上,假设物种之间彼此相互独立。多物种群落的时间稳定性Sm=(m1+m2+…+mn)/(σ2(x1)+σ2(x2)+σ2(x3)+…,σ2(xn))1/2,其中m为物种的多度或生物量,假设任何组成物种的变异系数与单物种群落的变异系数相同[33],即CVi=CVs. 100σ(xi)/mi=100σ(μ)/μ,σ(xi)2=(σ(μ)/μ)2mi2=(1/Ss)mi2,因此,多物种群落的时间稳定性可以表述为:
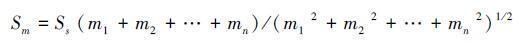 (4)
(4)
对群落中所有物种按多度的大小排序,多度递减程度由变量a决定。a越大说明群落的均匀度越小。假定mi=m1exp[-a(i-1)][33],则公式(3)可以写为:
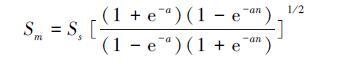 (5)
(5)
通过公式(5)可以发现,群落的时间稳定性随物种数n的增加而增加,同时受到均匀度的影响如图 3所示。
均匀度越高,则群落中每个物种的作用就越大,由于投资组合效应减少群落波动性的程度就越大[33]。随着均匀度降低(优势度增加)群落时间稳定性与物种数之间的正相互作用会受到破坏,最终,由于物种对群落的贡献非常的不均匀以致增加群落物种的数目将不会影响群落的时间稳定性[5](图 4)。此外降低均匀度会引起群落物种数的减少[54],因此会影响群落的生产力[45],进而间接地影响群落的时间稳定性。均匀度通常对人类活动和环境变化比丰富度更加敏感,也许在物种灭绝之前已经对系统过程产生影响[55-56],在生物多样性与生态系统稳定性的研究中,生态学家应该重视既影响群落稳定性又影响生物多样性的过程[53]。此外,Hillebrand等[56]综述了均匀度对稳定性的影响,发现均匀度的提高会提高恢复力,因为在受到破坏后,均匀度高的群落中最适宜的物种增加多度恢复生态系统功能所需的时间更短。
组成物种的稳定性高会使群落更加稳定[57]。物种的波动性小,则群落的方差之和会减少,进而减少群落的时间波动性。在自然系统中,系统的功能属性在短期内主要由优势种的性质决定,一些研究尤其是野外试验发现优势物种会限制生物多样性与稳定性的关系[58-59]。一些研究表明优势种比其他物种的稳定性更高[58, 60],增加稳定物种的相对多度会增加群落的稳定性[60]。
3.2.3 平均值-方差关系Taylor[61]发现在生物群落中,方差(σ2)和多度的平均值(μ)之间存在指数关系,即Taylor指数方程:
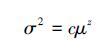 (6)
(6)
式中,c为常数,z为标定系数。
Tilman[8]在此基础上提出:
单物种群落的标准差为σs=c1/2μz/2,单物种群落的时间稳定性为:Ss=μ/σs=c-1/2μ1-z/2
假设(1)n个物种的群落总多度同样为μ,也就是物种总多度不随物种数改变
假设(2)群落中所有物种的多度相等,则每一个物种的多度为(μ/n)
假设(3)物种之间不存在相互作用
假设(4)n个物种的群落z值与常数c均与单物种群落相同
则n个物种的群落中每个物种多度的方差为σ2=c(μ/n)z,群落所有物种的方差之和为σn2=nc(μ/n)z,σn=c1/2μz/2n(1-z)/2,则n个物种群落的时间稳定性为: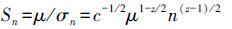
所以n个物种群落的时间稳定性与单物种群落的时间稳定性的比值为:
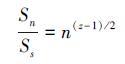
当z>1时,这一比值会随着多样性的增加而增加,也就是投资组合效应会对群落属性起稳定作用[52]。z值的范围会决定群落的稳定性和多样性之间的关系,在z>1后,z值越大,稳定性受物种多样性的影响就越大。将群落中所有物种的多度μ和方差σ2做对数转换,log(σ2)和log(μ)呈线性关系,其斜率便为标定系数z[58]。在给定多度下,z值越大,说明方差σ2随多度的变化越剧烈,因此在自然群落中,z值更直接的意义代表着群落组成物种的波动程度[60]。
3.3 协方差之和效应从公式(2)中可以推断,物种协方差之和发生变化会引起群落时间稳定性的改变。直接的,一些研究用物种的协方差之和来判断群落是否存在补偿动态[62],它是指在群落中,一个物种多度的增加伴随着其他物种多度的减少。这种负的相互作用会降低群落多度的协方差之和,因此被认为是维持群落稳定性机制之一。如果协方差之和为负值,则补偿动态在维持稳定性方面起重要作用[62]。协方差之和往往受竞争和物种响应的异步性的影响。在自然界中二者很难区分,例如在Tilman[48]的试验中,4种植物对干旱的响应与其他物种不同,但很难判断这种反应是因为种间竞争还是单纯对干旱的响应不同引起。生物多样性高的群落会有更大机率包含对环境响应不一致的物种,同时会使物种间的作用更加复杂,因此一些研究认为多样性高的群落,补偿动态应该更强[60]。
3.3.1 竞争群落中物种竞争有限的资源会使部分物种多度增加而另一些物种多度降低,因此竞争会使协方差之和变小,也就是使公式(2)中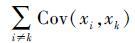 的值降低,一些研究认为竞争可能会增强群落的时间稳定性[8, 52]。然而Ives等[34]发现,竞争会增加种群波动的振幅,使物种波动更剧烈,所引起的减少协方差效应会通过增加方差效应所抵消,因此种间竞争对群落稳定性的影响不明显。在竞争对稳定性作用的研究中,部分生态学家曾用负的协方差之和来定量化物种间的竞争作用[63-64],事实上,如果两个物种负相关,第3个物种不可能与前两个物种之间均为负相关关系[46],在不存在较强的物种相互作用的情况下,一些物种相对多度的增加会被其他物种多度的减少所补偿,物种相对多度变化是一个总和相加为零的游戏[65],在自然群落中,由于物种经过生境条件的筛选往往对环境波动的响应比较相似,因此在许多群落中协方差之和更趋向于为正值[66]。因此竞争减少协方差之和的作用不会随着群落中物种数目的增加而增加[8],仅仅依靠负的协方差之和来判定竞争的作用并不合适[67-68]。最近,Loreau 与 de Mazancourt[7]用理论模型发现,与以前的研究结果不同,物种的竞争会使系统属性更加不稳定,因此增加系统稳定性需要减弱种间竞争的强度。
的值降低,一些研究认为竞争可能会增强群落的时间稳定性[8, 52]。然而Ives等[34]发现,竞争会增加种群波动的振幅,使物种波动更剧烈,所引起的减少协方差效应会通过增加方差效应所抵消,因此种间竞争对群落稳定性的影响不明显。在竞争对稳定性作用的研究中,部分生态学家曾用负的协方差之和来定量化物种间的竞争作用[63-64],事实上,如果两个物种负相关,第3个物种不可能与前两个物种之间均为负相关关系[46],在不存在较强的物种相互作用的情况下,一些物种相对多度的增加会被其他物种多度的减少所补偿,物种相对多度变化是一个总和相加为零的游戏[65],在自然群落中,由于物种经过生境条件的筛选往往对环境波动的响应比较相似,因此在许多群落中协方差之和更趋向于为正值[66]。因此竞争减少协方差之和的作用不会随着群落中物种数目的增加而增加[8],仅仅依靠负的协方差之和来判定竞争的作用并不合适[67-68]。最近,Loreau 与 de Mazancourt[7]用理论模型发现,与以前的研究结果不同,物种的竞争会使系统属性更加不稳定,因此增加系统稳定性需要减弱种间竞争的强度。
在自然界中,由于不同的物种具有不同的属性,因此对环境波动的响应不可能完全相同[67]。这种异步性一方面体现在对环境波动反应的方向相反。例如,一些物种在特定环境波动下多度增加,而另一些物种的多度减少,这种对环境波动的相反反应会造成群落总多度的波动减弱,进而增加群落的时间稳定性。另一方面体现在环境波动发生后的反应速度不同[7]。例如,一些物种在发生干扰后的t时间发生改变,而另一物种则在t+1时间内反应,因此可以产生时间生态位分化,从而使群落总多度的波动趋于平缓。理论模型研究认为该机制在维持群落稳定性方面的作用非常关键[7, 34, 40]。例如Ives等[34]强调了物种对环境波动的敏感性。最近的两个模型[7, 40]认为该机制可能是影响稳定性的主要机制。Hector等[46]在欧洲草地实验中对仅含两个物种的群落进行分析,发现两个物种生物量负的时间相关关系越强,群落的稳定性越高,证明物种反应的不同时性是影响群落时间稳定性的机制之一。但是关于这个机制的实验研究数量非常稀少,可能因为现有生物多样性实验开展的时间有限,限制了对该机制的实验验证。
4 生物多样性与稳定性研究中的争议在近半个世纪的生物多样性与稳定性研究中,无数的学者投入了无限的研究热情。然而研究数量的增加没有使生物多样性与稳定性研究的争论风暴平息,反而引出了更多悬而未决的议题,这也表明了该领域的复杂性。生物多样性与稳定性均具有十分广泛的定义,其中对稳定性的定义和对生物多样性的定义已有详细介绍[33, 42],不同的定义会得出不同的生物多样性稳定性关系[41-42]。任何事物的关系可以由方向和强度两方面概括,生物多样性与稳定性关系的方向仍存在争议,例如Ives与Carpenter[2]对52个研究进行分析,发现有69%的研究报道了生物多样性与稳定性正相互关系,14%的研究发现生物多样性与稳定性存在负相互关系,17%的研究未发现二者之间存在明显作用。在强度方面,影响稳定性的影响因子众多,而其他因子如非生物因子在影响稳定性的同时也会对生物多样性产生影响,从而使生物多样性对稳定性的影响更加难以预测。除此以外,生物多样性与稳定性研究的具体争议还包括以下几点。
4.1 不同机制对稳定性作用的争议(1) 诸多稳定性机制相互之间并不独立,一个机制的变化往往会引起其它机制的改变。例如,生态位互补会引起种间正相互作用,从而使群落的生产力增加,产生超产现象[51],影响物种对环境波动响应的异步性,同时还会减弱物种对资源的竞争强度,从而使其对生态系统稳定性的作用更加难以评定。均匀度本身会影响系统的稳定性,而随着均匀度的改变投资组合效应在维持系统稳定性方面的作用强度会受到影响[5],当均匀度低至某一下限后,群落稳定性仅仅会受到优势种稳定性的影响而不会受到生物多样性的影响。
(2) 一些机制在引起稳定性增加的同时,还具有使稳定性降低的作用,因此同一机制有可能引起不同的生物多样性稳定性关系[2]。竞争对稳定性的影响一直备受争议[7, 33-34],在前文已经有所介绍这里不做赘述。生态位互补效应会引起超产[51],因此对稳定性的提高具有促进作用,但另一方面,由其引起的种间正相互作用会使协方差之和增加,从而会使群落多度的时间波动性加剧。该机制是否对稳定性起促进作用有赖于其引起的超产效应和增加协方差之和效应的作用强度。
(3) 不同的机制在不同的研究中作用强度不统一。一项整合研究发现超产比方差之和与协方差之和的作用明显[10]。波动环境中随机群落动态模型的研究则强调物种对环境波动响应的异步性[7, 34, 67]。而在自然群落中,很多研究发现优势种的稳定性对群落的稳定性影响很大[58-59]。不同的实验研究在不同区域开展,巨大的地理环境差异造成不同实验的群落组成相差甚远,然而稳定性机制如投资组合效应、均匀度、优势种的稳定性、竞争、对环境波动响应的异步性等均受到群落结构的影响,这使得在生物多样性与稳定性研究中各机制的作用强度存在很大争议。
(4) 统计学机制和生物学机制间的争议。在投资组合效应提出以前,生态学家往往忽视了统计学机制的作用而多关注种间相互作用。Doak等[33]认为统计平均效应在维持生物多样性稳定性关系中必然会发挥作用,即使在前人已经报道的竞争群落中,统计平均在短期内发挥的作用有可能强于竞争的作用。Tilman等[52]表明投资组合效应影响生物多样性稳定性关系与否还取决于标定系数z的取值,也就是投资组合效应会受到群落自身性质的影响。统计学机制不可避免的会受到生物学机制的影响,在自然群落中统计学机制的作用强度一直深深困扰着生态学家。也许将统计学机制作为零和假设会为生态学家研究物种相互作用的影响提供帮助。
4.2 生物多样性与稳定性研究方法的争议在生物多样性与稳定性关系研究中,众多研究方法主要可分为理论模型研究和实验研究两类。de Mazancourt等[40]将主要的模型方法分为四类:(1) 统计方法,如Doak等[33]对投资组合效应的推演方法;(2)随机动态方法,描述种群多度对环境波动的响应,但不考虑种间相互作用;(3) 一般的种群动态方法,既考虑种群动态对环境波动的响应,又考虑种间相互作用;(4)关于种间竞争的特定模型,其中反馈机制决定着物种的共存。实验方法同样多种多样,大体可分为实验室试验和野外实验。野外实验包括大尺度的自然实验和设计精细的样地水平的实验[69-70],同样还可分为控制实验和观测实验。即使是控制实验仍然可以分为对生物多样性(丰富度)进行直接控制的实验和非直接控制的实验[11-12]。各种不同的研究方法不可避免的会造成理论研究与实验研究的矛盾,理论研究间结论的不一致,实验结果间的不同[2]。
理论和实验结果间存在矛盾[71]。首先,在进行理论研究中往往需要提出理想假设,而这些模型在自然界中很难满足。如前面提出的4个假设,假设1为几个物种的总多度或总的生产力不随物种改变。但是生物多样性与生产力间存在正关系[45],以及不存在关系的情况在自然界中都广泛存在[60]。假设2为群落中所有物种的总多度相等。但往往不同的群落具有不同的群落结构,因此该假设在自然界中很难实现,在野外实验中,许多研究表明比例假说(mass-ratio hypothesis)往往是比较重要的假说[58-59, 72]。假设3为物种间不存在相互作用。在自然界中互利共生和竞争广泛存在,且种间的正相互作用和竞争还会随着环境梯度的改变发生转换[73]。假设4为n个物种的群落z值与常数c均与单物种群落相同。但是z值往往通过拟合得来,与物种组成和物种的波动密切相关,因此变化更加复杂。其次,许多理论研究和实验类研究应用的稳定性概念不同,这也许是造成20世纪70年代对生物多样性与稳定性关系研究与实验类研究结果不同的主要原因之一[5]。例如May[74]曾定义稳定性为在受到干扰后返回多物种平衡状态的能力,而大多数的实验研究则主要关注群落多度的变异性。
20世纪90年代的一些生物多样性实验的兴起,开创了生物多样性与稳定性研究的一个新时代。然而对这些实验的解释引起了广泛的争议,其中对生态箱实验、Cedar Creek草地多样性试验、微宇宙实验以及欧洲草地实验的有关争议已有详细的综述[30]。随后人们开始将争议的矛头指向了实验设计方法的本身[14]。控制生物多样性的实验大多发现了生物多样性与稳定性的正相互关系[11],而在自然群落的观察研究发现了生物多样性与稳定性之间存在其他形式的关系[69]。观察研究认为控制实验被限制在有限的区域内,其结果与非实验系统的结果存在差异,这些限制了生态学家对这些结果进行归纳总结并提出可靠地预测[12]。一些生态学家试图解释控制实验和观测实验结果的不同,认为在自然群落空间的异质性限制了生物多样性与稳定性关系的体现,同时实验系统的群落与自然群落存在本质上的差异也是重要的原因之一[10, 75]。
4.3 生物多样性与物种水平和群落水平的稳定性关系不一致May在1973年提出群落的属性比种群的属性更加稳定[26],这一观点在当时一直被忽略。随后的一些实验研究和理论研究表明生物多样性可能会增加群落水平的稳定性而降低种群的稳定性[8, 43, 45-46, 48]。但最近的一些研究表明,在一些特殊情况下生物多样性有可能既提高群落的稳定性又提高种群的稳定性[10, 76-77]。Campbell 等[32]对已发表的35个研究进行整合分析,结果表明生物多样性对群落稳定性有增加效应,但在36个生物多样性与种群稳定性的关系中,19个具有负相关关系,17个正相关关系,使得生物多样性对种群稳定性的影响更具争议。生物多样性与物种水平和群落水平的稳定性关系不一致与研究所选取的系统有关:生物多样性与稳定性研究在草地等陆地生态系统开展最多,还有一些研究在水生生态系统展开,此外还有一些研究关注微生物群落的稳定性[2]。其中陆地生态系统通常仅包含一个营养级,而水生生态系统往往会选择多个营养级。Jiang与Pu[10]表明在单营养级群落中,生物多样性会增加群落稳定性降低种群水平稳定性,而在多营养级群落,生物多样性对群落稳定性和种群稳定性均有提高的效应。
5 结语近年来,随着野外实验开展时间的增加,许多实验有条件对稳定性进行研究,因此关于稳定性的研究数量一直持续增加。国外著名的实验仍在持续对生物多样性和稳定性关系及其内在机制进行探索。例如Tilman[45]等在Cedar creek草地实验中发现生物多样性与稳定性的正相互作用,而在作用机制方面该研究强调了投机组合效应和超产。Hector[46]等发现在欧洲草地试验中生物多样性提高了群落稳定而抑制了种群水平的稳定性,超产和种群波动的异步性是该实验中的重要稳定性机制。国内的相关研究也取得了一定的进展,如Bai等[78]通过对内蒙古典型草原的长期观测数据的分析同样发现了生物多样性对群落稳定的促进作用和对种群稳定性的抑制作用,同时该研究对补偿动态进行了探讨。Yang等[79]对7a的野外数据进行分析表明割草增加了群落水平和种群水平的稳定性,而施肥的效果恰好相反。在理论研究方面,生态学家正在试图建立更加综合的理论,并继续对以前的机制进行推演[7, 71]。新的理论更加关注物种的内在性质,同时将理论结果与实验结果相结合对诸多的稳定性机制进行检验[40]。
从最近的研究中我们可以总结一些未来实验研究的可能趋势。(1) 更注重长期野外实验的研究。如Reich等[17]对两个超过13a的生物多样性实验分析发现,生物多样性的作用强度会随着试验时间的增加而增加。(2)对多个野外实验进行综合分析加深认识。如Isbell等[16] 综合分析了17个生物多样性实验的结果,Tilman等[9]对11个实验进行整合分析,而Hooper等[13]的研究中综合了16个实验的结果。(3) 整合分析方法的运用,由于条件的限制,许多生态学家往往只能获得有限实验的原始数据,对已发表的研究进行整合分析提供了新的思路。Cardinale等[50]人的整合分析对选择效应和互补效应对生物多样性实验的解释效果进行了比较,Jiang与Pu[10]则通过整合分析的方法比较了几个稳定性机制的作用强度。因此,增加实验研究的时间尺度和空间尺度,结合整合分析的方法有可能为我们理清诸多稳定性机制之间的关系和相对强度提供途径,对作用强度最大的机制加强研究将会有助于提高理论认识。理论的发展将会加深我们对生物多样性稳定性的关系认识,预测生物多样性丧失对群落稳定性的影响,以及用来指导未来生物多样性的实验研究[7, 38]。
同样,将来的研究应多关注非多度/生物量的属性[10],控制实验应该控制具有不同属性物种的多度而不要只单纯的控制物种数目。现在关于时间稳定性概念的应用多是关注多度或生物量,用其他变量的变异性定义稳定性后,现有的稳定性机制是否仍会发挥作用或者作用强度是否会发生改变需要生态学家进行探索。这方面的工作已经开展,如最近一项在内蒙古典型草原的稳定性研究应用丰富度和盖度的变异性作为稳定性的定义[79]。未来的研究可以关注均匀度的时间变异性,因为它比丰富度的变化更加敏感,同时对丰富度和稳定性均有影响[56]。此外,现在关于如何标准的测量植物属性投入大量的努力并且取得了一定的发展[80-81],这些努力使得全球很多区域的植物属性得以测量[82-84]。这些属性包括对环境波动的响应速度,以及物种的稳定性,通过控制具有特定属性物种的多度,可以使人们进一步理解和预测生物多样性对稳定性的影响[7]。
呼吁加强对生物多样性的保护意识,采取对生物多样性的保护措施。生态学家通过设计实验为政策和管理提供启示[32]。在实验中得出的新理论可以对生物多样性丧失后的可能结果进行预测。然而从实验的设计到得到可信的实验结果,再到理论的提出,往往会花费很长的时间。由于政策和科学间的鸿沟,根据理论制定合理的管理措施可能比较困难,而从管理措施的实施到这些管理措施发挥作用又需要很长的时间[14]。然而生物多样性在全球尺度的丧失速度从未减缓,据报道在未来的240年间,全球物种有可能会经历第六次大灭绝[85]。此外,很多物种在提供特定的生态系统功能和服务方面具有重要作用,尚有很多物种的潜在作用还不能被人们很好的理解。因此,为了不至发生"无米之炊"的悲剧,提高人们对生物多样性保护的意识和采取对生物多样性的有效保护措施迫在眉睫。